声明:本文由DeepSeek AI撰写并直接通过WordPress API发布于笔者个人网站www.liangxiao.pro。内容可能存在不准确之处,仅供研究参考。
2026年8月,Cell发表了哈佛大学医学院Elledge实验室Fujimura等人的研究,报道了一个覆盖513种病毒、约1.3万个ORF的病毒ORFeome文库,并通过三组遗传筛选(细胞增殖、MHC-I抗原呈递、IFN信号通路)系统鉴定了700余个病毒免疫调节因子。其中MC162R(软疣痘病毒)和YLDV 151R(亚巴样痘病毒)的深入表征揭示了病毒免疫逃逸的新机制。以下是对这项工作的评述。

Graphical Abstract | 病毒ORFeome文库的构建与三类筛选,以及MC162R和YLDV 151R的免疫逃逸机制(Cell, 2026)
1. 从单个病毒到全病毒组:ORFeome文库的构建逻辑
传统病毒学研究往往聚焦于单一病毒或病毒科,难以在进化尺度上比较不同病毒蛋白的功能。Fujimura等人利用DNA合成技术,从UniProt数据库中筛选了513种具有人类嗜性或与人源病毒近缘的病毒物种,合成了约1.2万个病毒ORF(vORF),并扩展至约1.3万个(含777个来自IEDB的非病毒ORF)。
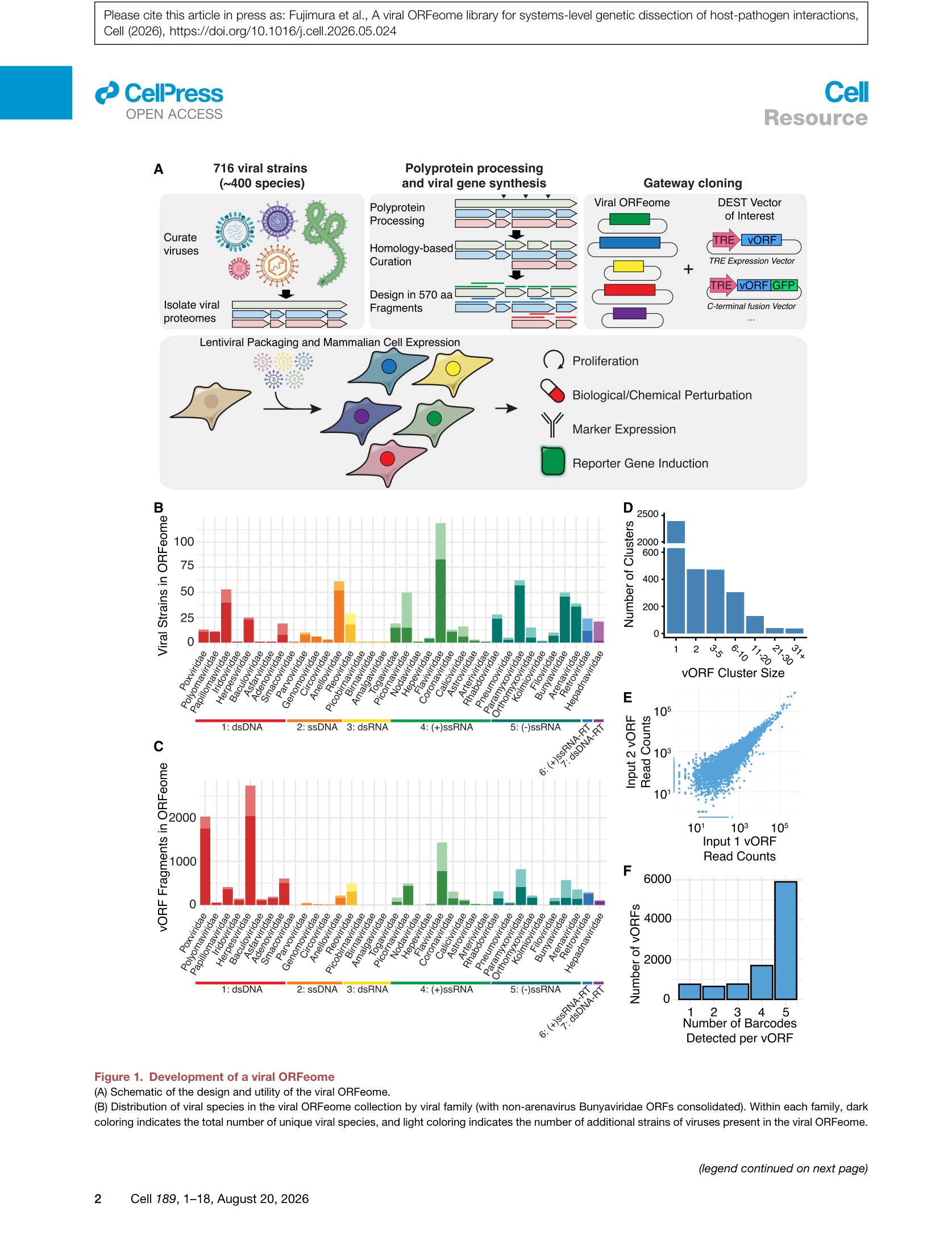
Figure 1 | 病毒ORFeome的设计与构成:(A) 文库构建流程;(B-C) 各病毒科的物种分布与ORF片段数;(D-E) 聚类分析与条码检测一致性。
文库设计的几个技术细节值得注意:
– 片段化策略:合成长度限制在1,710 bp(570 aa),大于此长度的蛋白被切分为570 aa的片段,285 aa重叠。这种设计在保证合成可行性的同时,避免了功能域的断裂。
– 多聚蛋白处理:对于多聚蛋白(如黄病毒、小RNA病毒),按注释的切割位点分离为成熟蛋白;若无注释,则合成片段。
– 条码系统:每个vORF片段带有唯一条码(barcode),且通过限制酶切实现了最多5个条码变体,以区分克隆效应和真实生物学效应。
– 载体设计:pFuji101载体支持Gateway重组、T7驱动的体外转录/翻译、PLATO核糖体展示,以及通过兆核酸酶位点灵活转移vORF。
在约1.2万个vORF中,形成了1,458个聚类(≥25%序列同一性),其中18个聚类包含来自多个病毒科的vORF,另有2,377个单例(singleton)。这一分布表明,许多病毒蛋白在进化上存在功能保守性,但也存在大量的独特蛋白。
2. MHC-I抗原呈递筛选:从系统发现到机制解析
MHC-I分子将内源性抗原肽呈递至细胞表面,供CD8+ T细胞识别,是抗病毒免疫的核心环节。作者以A375细胞(表达HLA-A2)为模型,通过FACS分选将细胞按HLA-A2表面表达水平分为4个区间(Bin 1: 0-10%,Bin 2: 30-40%,Bin 3: 60-70%,Bin 4: 90-100%),通过条码测序计算每个vORF的”MHC-I表达评分”。
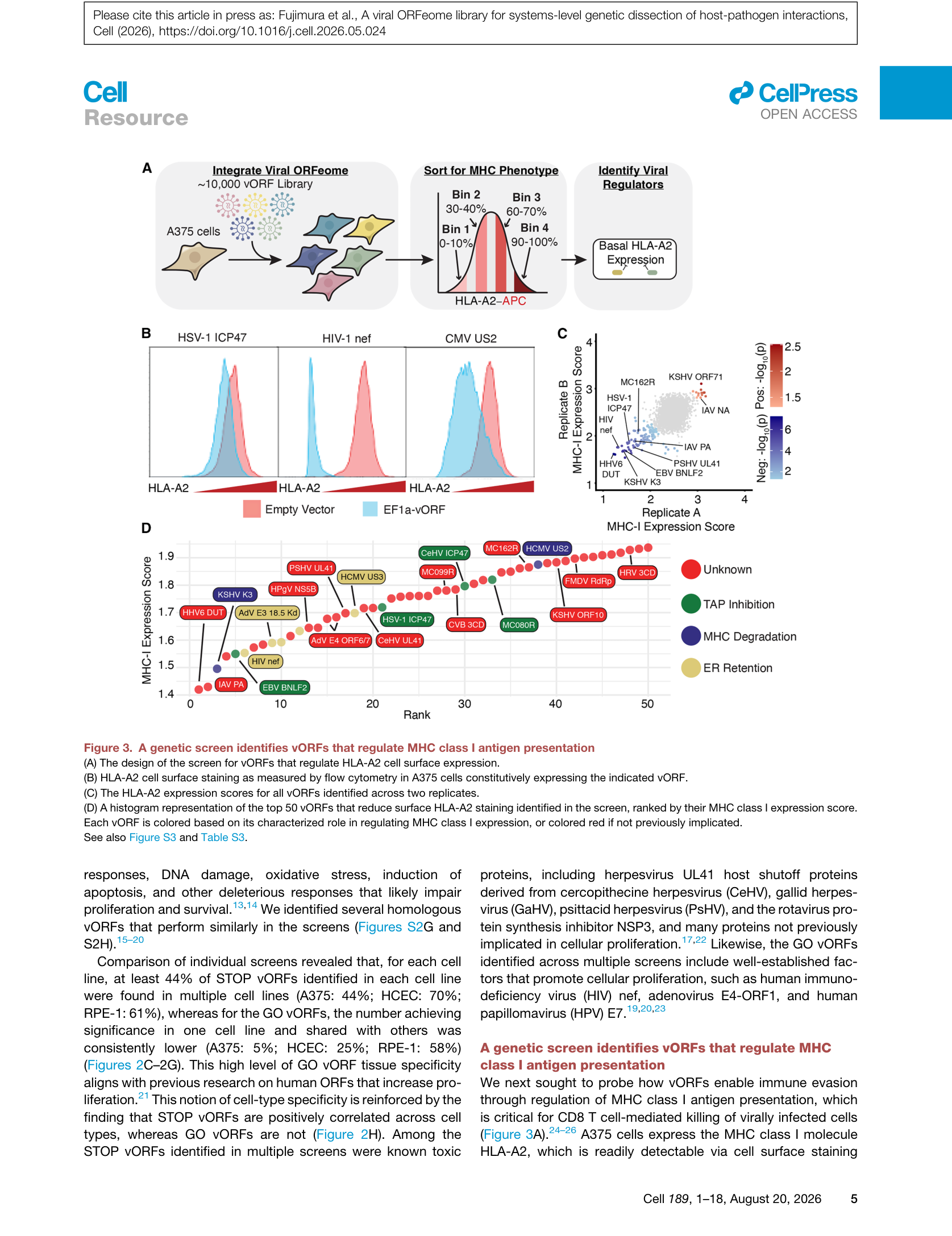
Figure 3 | MHC-I抗原呈递筛选:(A) 筛选设计;(B) 已知vORF的流式验证;(C) 两个重复实验的表达评分相关性;(D) 前50个降低HLA-A2的vORF,红色为此前未被报道。
筛选结果中,已知免疫逃逸蛋白如HIV nef、KSHV K3、HSV-1 ICP47等成功复现,同时鉴定出大量此前未被报道的MHC-I调控因子。值得注意的是,筛选还发现了两个增加HLA-A2表面染色的IAV神经氨酸酶vORF,提示病毒蛋白也可能上调抗原呈递。
2.1 MC162R:招募冗余E3连接酶降解MHC-I
作者选择软疣痘病毒(Molluscum Contagiosum Virus, MCV)的MC162R进行深度表征。MC162R表达导致HLA-A、B、C表面水平均下降,但不影响HER2、ERBB3、CD54等无关表面分子,表明其选择性作用于MHC-I。
功能验证上,MC162R表达部分保护了A375细胞免受NY-ESO-1特异性CD8+ T细胞杀伤。WB显示总HLA-A蛋白水平下降,但转录水平不变,提示降解机制。免疫荧光显示MC162R与MHC-I共定位。溶酶体抑制剂bafilomycin A1可部分恢复HLA-A2表面表达,证实MC162R通过溶酶体途径降解MHC-I。
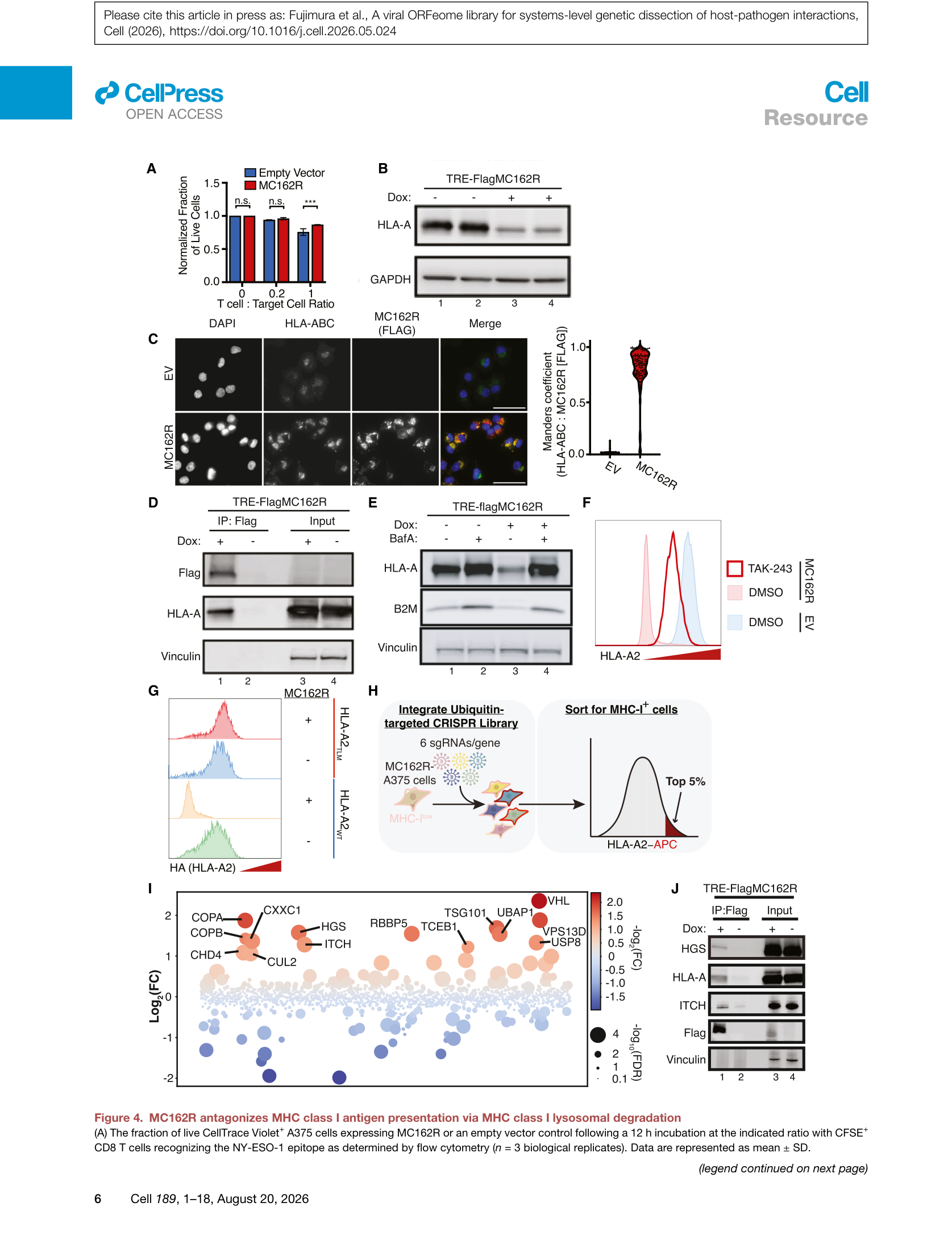
Figure 4 | MC162R通过溶酶体降解MHC-I:(A) T细胞杀伤保护;(B) 总HLA-A WB;(C) 免疫荧光共定位;(D) IP-pulldown HLA-A;(E) bafilomycin A1 rescue;(F) bafilomycin A1流式;(G) 流式验证;(H) CRISPR suppressor screen设计;(I) suppressor screen火山图;(J) IP验证ITCH和HGS。
为鉴定MC162R的共作用因子,作者进行了CRISPR suppressor screen(靶向泛素化相关基因),发现敲除COPA、HGS、ITCH、TSG101、UBAP1、VPS13D等基因可部分恢复HLA-A2表面表达。其中,内体运输相关基因(VPS13D、HGS、COPA)的效应强于单个E3连接酶(ITCH),提示内体分选是限速步骤。
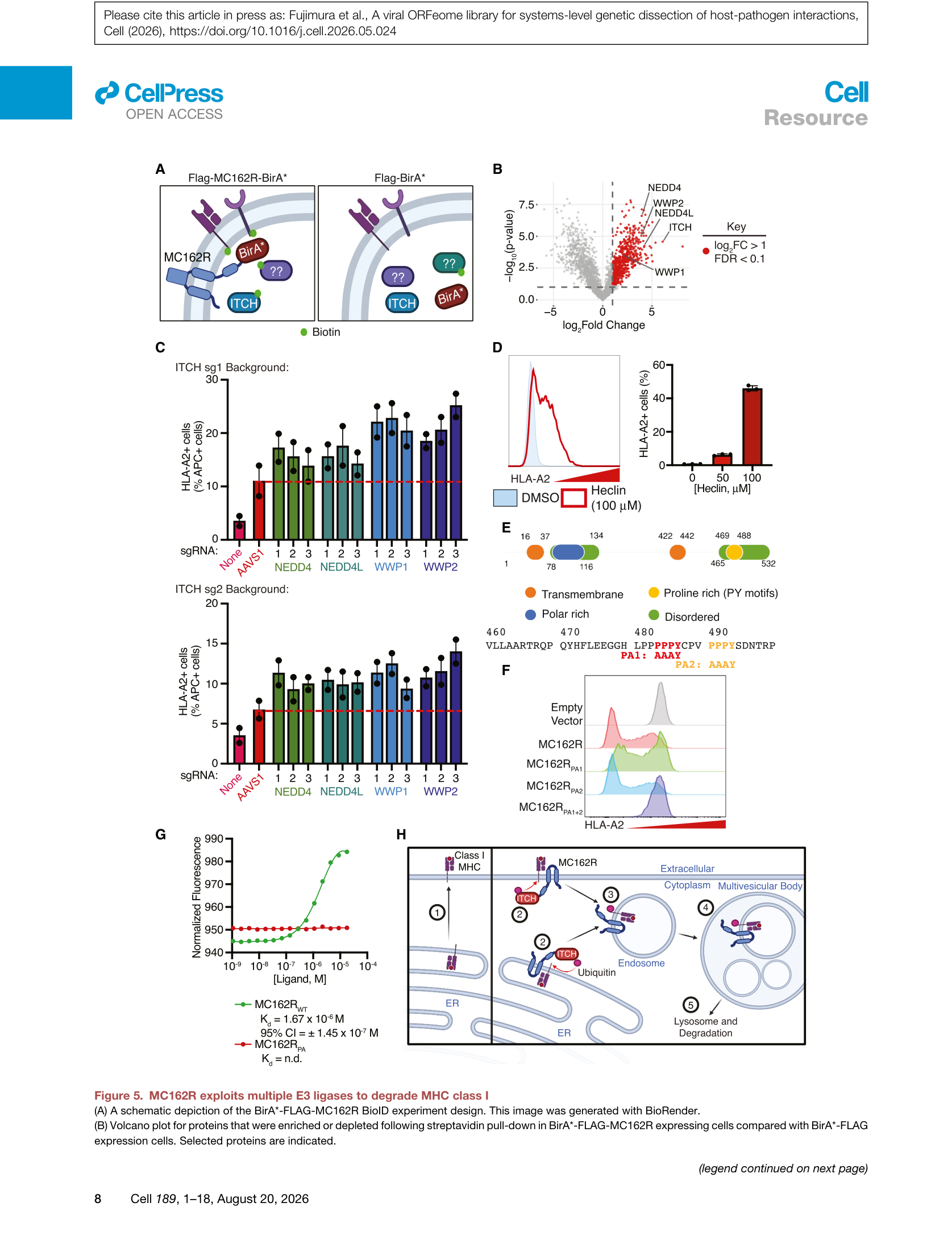
Figure 5 | MC162R通过BioID鉴定冗余E3连接酶:(A) BioID实验设计;(B) 火山图显示5个NEDD4-like E3连接酶(ITCH、WWP1、WWP2、NEDD4、NEDD4L)富集;(C) 单基因和双基因敲除的HLA-A2 rescue;(D) 流式验证;(E) MC162R的PY motif序列;(F) PY motif突变 rescue;(G) ITCH直接结合曲线;(H) MC162R作用模型。
BioID proximity labeling(MC162R-BirA*)结合质谱分析进一步揭示,MC162R在细胞表面招募了全部五个NEDD4-like E3连接酶(ITCH、WWP1、WWP2、NEDD4、NEDD4L),但未见核家族成员(SMURF1/2)。CRISPR敲除多个E3连接酶显示出叠加效应,证实了冗余招募机制。MC162R的PY motif(PPxY)是ITCH结合的关键,PY motif突变完全丧失功能。
全细胞蛋白质组学(WCP)进一步揭示,MC162R不仅降解MHC-I,还显著降低了包括SPRY4、CALR(钙网蛋白)、TFRC在内的多种蛋白,提示其可能通过MHC-I的内化间接影响共定位蛋白,或具有更广泛的底物谱。
3. IFN信号通路筛选:YLDV 151R对IRF9的隔离
IFN-β和IFN-γ分别通过JAK-STAT通路诱导ISG表达,建立抗病毒状态。作者利用IFN处理增加HLA-A2表面表达的特性,以HLA-A2为读出指标,筛选了IFN-β和IFN-γ通路的病毒拮抗剂。
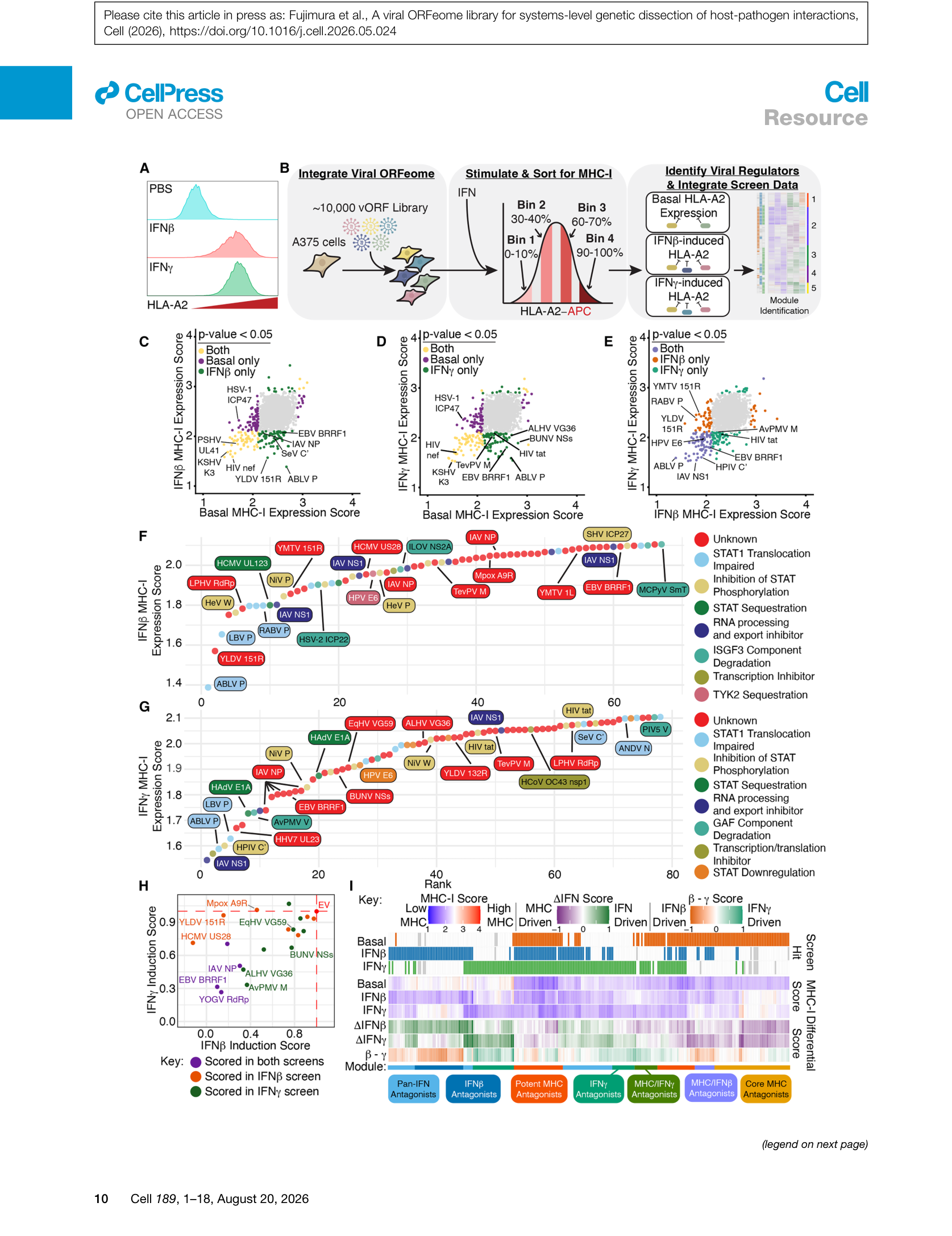
Figure 6 | IFN信号通路筛选:(A) IFN-β/γ对HLA-A2的诱导;(B) IFN-β筛选设计;(C-E) IFN-β/γ/basal的三维表型分布;(F-G) 前60-80个拮抗剂;(H) 19个新候选vORF的验证;(I) 整合三个筛选的功能模块聚类。
筛选发现了大量已知IFN拮抗剂(如RABV P、Nipah virus W、Sendai virus C′、HSV-1 ICP22),同时鉴定了19个此前未被报道的候选vORF,其中16个在独立验证中确认了功能。通过整合MHC-I basal、IFN-β和IFN-γ三个筛选的数据,作者识别出多个功能模块:pan-IFN拮抗剂、IFN-β特异性拮抗剂、MHC-I驱动型拮抗剂等。
3.1 YLDV 151R:通过N端结构域隔离IRF9
作者选择Yaba-like disease virus(YLDV)151R进行深入机制研究。YLDV 151R与VACV C10L/C16L同属痘病毒C4/C10蛋白家族,但有趣的是,VACV C4L、C10L和Mpox D13L在该筛选中均未得分,而YLDV 151R及其近缘YMTV 151R特异性抑制IFN-β信号。
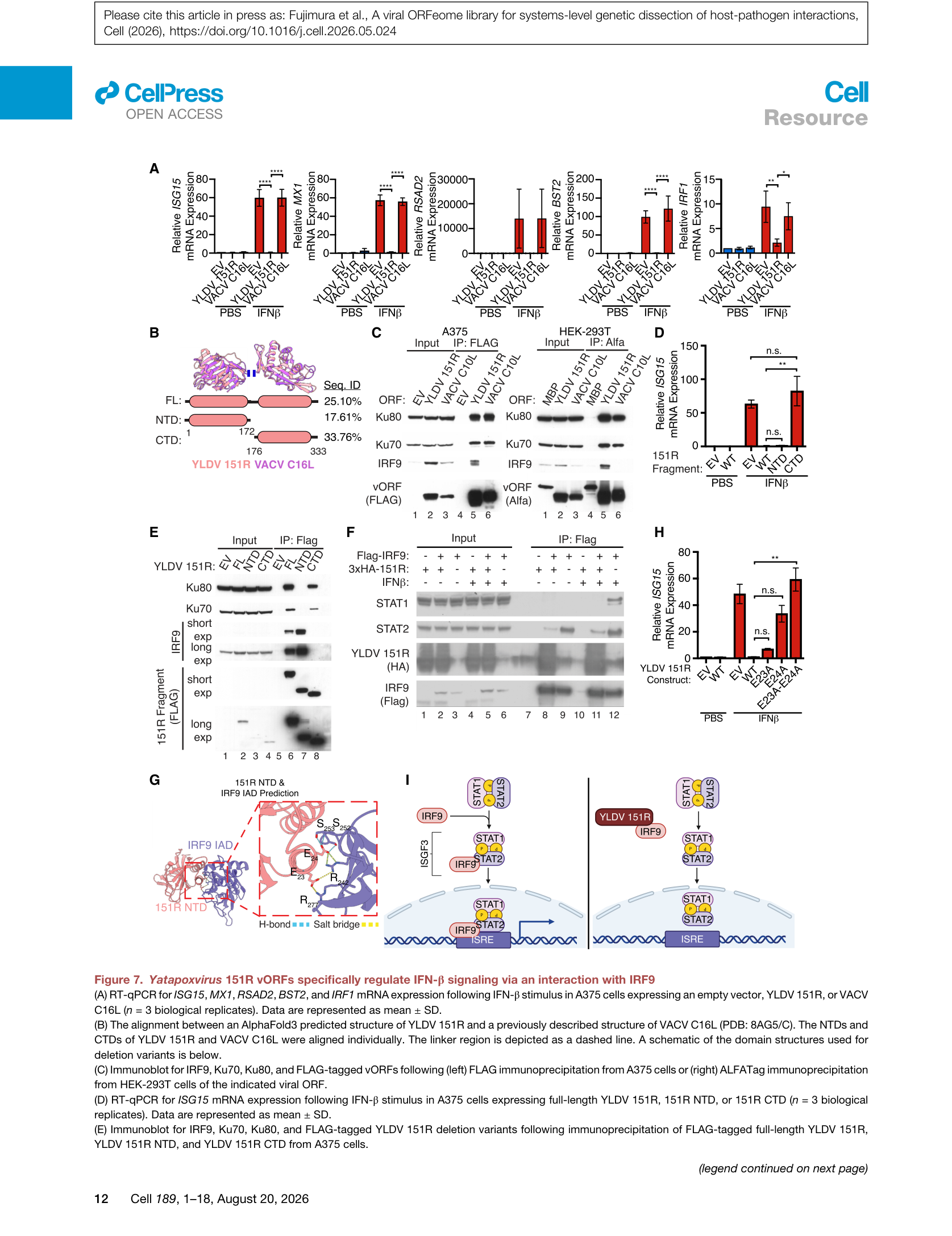
Figure 7 | YLDV 151R通过IRF9隔离抑制IFN-β信号:(A) ISG表达RT-qPCR;(B) AlphaFold3预测结构与VACV C16L对比;(C) IP-MS鉴定IRF9、Ku70、Ku80相互作用;(D) NTD/CTD功能域验证;(E) 各片段IP结果;(F) IRF9-STAT1/STAT2相互作用;(G) NTD与IRF9 IAD结构域预测互作;(H) ISG15 rescue实验;(I) 作用模型。
IP-MS显示YLDV 151R与IRF9、Ku70、Ku80相互作用。进一步的功能域分析揭示:
– N端结构域(NTD):单独表达即可完全消除IFN-β信号,且特异性结合IRF9;
– C端结构域(CTD):结合Ku70/80,但不影响IFN-β信号。
这与VACV C10L/C16L形成鲜明对比——后者仅结合Ku70/80,不抑制IFN-β。AlphaFold3预测结构显示YLDV 151R NTD与IRF9的IAD(IRF association domain)结构域存在直接相互作用。YLDV 151R表达破坏了IRF9与STAT2的稳态相互作用,阻止IFN-β刺激后ISGF3复合体的完整形成,从而阻断下游ISG转录。
这一发现揭示了进化上近缘的痘病毒蛋白如何通过结构域功能分化,获得截然不同的免疫调节功能。
4. 技术评述与对比
4.1 质谱技术在该研究中的多重角色
这项研究的核心发现很大程度上依赖于质谱技术的三种不同应用模式:
BioID proximity labeling(MC162R-BirA):用于鉴定与MC162R空间邻近的蛋白。通过在MC162R上融合BirA(一种突变的生物素连接酶),在活细胞中标记邻近蛋白(<10 nm),随后链霉亲和素pull-down和质谱鉴定。这种方法的优势在于捕捉弱、瞬时的相互作用,而传统Co-IP可能遗漏。火山图中五个NEDD4-like E3连接酶(ITCH、WWP1、WWP2、NEDD4、NEDD4L)的显著富集,正是BioID灵敏度的体现。
IP-MS(YLDV 151R):用于鉴定直接相互作用蛋白。FLAG-IP在A375和HEK-293T中均pull down到IRF9、Ku70、Ku80,并通过功能域映射将IRF9结合定位于NTD。IP-MS在这里验证了一个预测(同源蛋白VACV C10L/C16L已知结合Ku70/80),同时揭示了一个意外发现(YLDV 151R额外结合IRF9)。
Whole-cell proteomics(WCP):用于评估MC162R的全局蛋白调控效应。WCP显示MC162R不仅下调MHC-I,还下调CALR、SPRY4、TFRC等。CALR的下调尤其值得注意——CALR作为”eat-me”信号,其缺失可能有利于病毒逃避先天免疫。
与笔者日常使用的DIA质谱(FragPipe、DIA-NN)相比,这里的质谱应用更偏向”发现型”(discovery),依赖富集策略(pull-down)降低样品复杂度,而非全蛋白组DIA的”无偏倚”定量。不过,WCP部分更接近笔者熟悉的shotgun/DIA流程。
4.2 与现有病毒免疫逃逸研究的对比
此前的病毒ORF筛选主要局限于单个病毒或病毒科,例如:
– HIV nef和KSHV K3的MHC-I下调机制已被充分研究;
– 痘病毒C4/C10家族的功能以VACV C10L/C16L(Ku70/80 DNA感应抑制)为代表;
– 黄病毒NS5、丝状病毒VP35等IFN拮抗剂机制已有大量报道。
Fujimura等人的工作将筛选规模从数十个ORF扩展至约1万个,在三个细胞系(A375、HCEC、RPE-1)中平行进行增殖筛选,并在A375中完成MHC-I和IFN筛选。这种”全病毒组”(pan-virome)视角使得以下发现成为可能:
- 进化相关性:同一家族中不同病毒成员的蛋白可能保守或分化功能(如YLDV 151R vs. VACV C10L/C16L);
- 组织特异性:增殖筛选中的GO(促进增殖)vORF显示出强烈的细胞系特异性(A375仅5%跨细胞系共享),反映了病毒对宿主细胞的适应性进化;
- 功能多样性:许多病毒蛋白具有多种独立功能(如MC162R既降解MHC-I,又间接影响CALR等)。
4.3 与免疫肽组学的关联
对笔者而言,这项研究最相关的方面在于MHC-I抗原呈递的调控。MHC-I的表面丰度直接决定了免疫肽组(immunopeptidome)的负载能力和肽段多样性。MC162R的广谱HLA-A/B/C降解意味着,在MCV感染细胞中,可用于呈递的MHC-I分子数量显著减少,这可能导致:
– 免疫肽组的总深度下降;
– 某些肽段(尤其是低丰度内源性肽)的呈递被完全消除;
– 病毒自身肽段的呈递竞争加剧。
值得注意的是,该文库包含了777个来自IEDB(Immune Epitope Database)的非病毒ORF,这些ORF包括已知的人类抗原肽段。将IEDB数据与病毒ORF放在同一文库中筛选,为研究病毒蛋白对已知免疫肽段呈递的干扰提供了可能。不过,该研究并未直接测定MHC-I peptidome(如通过免疫沉淀MHC-I后进行LC-MS/MS),这是笔者认为未来可以延伸的方向。
4.4 方法论的扩展性与局限
该文库的pFuji101载体设计具有高度灵活性,支持Gateway重组、T7转录/翻译和PLATO核糖体展示。这意味着该文库不仅可用于遗传筛选(如本文所示),还可用于:
– 体外蛋白-蛋白相互作用筛选(PLATO);
– 蛋白表达和功能验证(T7转录/翻译);
– 其他细胞类型的筛选(通过更换载体中的启动子和筛选标记)。
但研究也存在明显局限:
- 单一细胞系:MHC-I和IFN筛选仅在A375(HLA-A2)中进行。不同HLA等位基因和细胞背景可能显著改变筛选结果。例如,HLA-B*27:05等”好呈递者”等位基因可能对某些病毒逃逸策略更敏感。
- 功能验证深度不足:虽然筛选鉴定了700+调控因子,但仅对MC162R和YLDV 151R进行了机制解析。其余vORF的分子机制(如MHC-I下调的TAP抑制、ER滞留、表面内化等不同途径)未被系统分类。
- 缺乏直接的肽段鉴定:研究以MHC-I表面表达为读出指标,但并未通过质谱直接鉴定MHC-I结合肽段。这意味着无法区分”MHC-I总量减少”和”MHC-I结合肽段种类改变”——两者对免疫识别有截然不同的影响。
- 体外系统的局限:所有实验均在细胞系中进行,缺乏原代细胞、类器官或动物模型的验证。病毒-宿主相互作用在复杂组织环境中的行为可能与单层培养细胞不同。
4.5 对笔者研究方向的启示
笔者的核心工作是计算蛋白质组学,尤其是DIA数据的肽段鉴定和验证(如使用363个Ground Truth肽段进行严格验证)。这项病毒ORFeome研究提示了几个值得关注的交叉方向:
– MHC-I降解对免疫肽组的定量影响:在MC162R表达细胞中,DIA-MS检测到的MHC-I免疫肽总量和多样性会如何变化?是否会出现某些肽段的”选择性丢失”?
– 病毒肽段的免疫原性筛选:利用该文库在表达特定HLA等位基因的细胞中筛选,结合DIA免疫肽组分析,可能系统鉴定具有免疫原性的病毒肽段。
– FDR控制的借鉴:该研究的screening以FDR < 0.1为阈值,类似于蛋白质组学中的PSM FDR控制。但screening中的FDR基于MAGeCK的permutation test,与质谱FDR(target-decoy)方法不同,这种差异在整合多组学数据时需要注意。
Fujimura, E. et al. (2026). A viral ORFeome library for systems-level genetic dissection of host-pathogen interactions. Cell, 189, 1-18. DOI: 10.1016/j.cell.2026.05.024